2 Multisensory mechanisms underlying the sense of the body and self
How the body shapes human conscious experience is an old and controversial philosophical debate. Yet, recent theories converge on the importance of sensory and motor bodily signals for the experience of a coherent sense of self and hence for self-consciousness in general (Berlucchi & Aglioti 2010; Bermúdez 1998; Blanke & Metzinger 2009; Carruthers 2008; Gallagher 2000; Legrand 2007; Metzinger 2007; Tsakiris 2010). Even the emergence of self-consciousness in infants has been linked to their ability to progressively detect intermodal congruence (e.g., Bahrick & Watson 1985; Filippetti et al. 2013; Rochat 1998).[2] The assumption that multisensory integration of bodily signals underpins the sense of a bodily self has opened up—next to clinical research—a broad and exciting avenue of experimental investigations in psychology and cognitive neuroscience as well as interdisciplinary projects integrating philosophy and neuroscience. Experiments in these fields typically provided participants with conflicting information about certain aspects of their body and assessed how it affected implicit and explicit aspects of the body and self. The first anecdotal evidence of an altered sense of self through exposure to a multisensory conflict dates back at least to the nineteenth century with the work of Stratton (1899). More systematic, well-controlled paradigms from experimental psychology have gained tremendous influence since the first description of the rubber hand illusion seventeen years ago (Botvinick & Cohen 1998). Since then, different important components underlying the bodily self have been identified, described, and experimentally modified. Most prominently: self-location—the feeling of being situated at a single location in space; first-person perspective—the centeredness of the subjective multidimensional and multimodal experiential space upon one’s own body (Vogeley & Fink 2003); body ownership—the sense of ownership of the body (Blanke & Metzinger 2009; Serino et al. 2013); and agency—the sense of being the agent of one’s own actions (Jeannerod 2006). In this section, we briefly describe these components of the bodily self as well as experimental paradigms that allow their systematic manipulation and investigation of their underlying neural mechanisms. Later, in section 4, we will describe how and to what extent vestibular signals might influence these components as well as their underlying multisensory integration.
2.1 Ownership, self-location, and the first-person perspective
2.1.1 Body part illusions
Both ownership and self-location[3] have traditionally been investigated in healthy participants using the rubber hand illusion paradigm (Botvinick & Cohen 1998). Synchronous stroking of a hidden real hand and a seen fake hand in front of a participant causes the fake hand to be self-attributed (i.e., quantifiable subjective change in ownership) and the real hand to be mis-localized towards the rubber hand (i.e., objectively quantifiable change in self-location). During the last ten years, various other correlates of the illusion have been described. For example, illusory ownership for a rubber hand is accompanied by a reduction of the skin temperature of the real hand (Moseley et al. 2008), an increased skin conductance and activity in pain-related neural networks in response to a threat toward the rubber hand (Armel & Ramachandran 2003; Ehrsson et al. 2007), and increased immune response to histamine applied on the skin of the real hand (Barnsley et al. 2011). Several variants of the illusion have been established using conflicts between tactile and proprioceptive information,[4] between visual and nociceptive information (Capelari et al. 2009), between visual and interoceptive information, and between visual and motor information (Tsakiris et al. 2007). All these multisensory manipulations have in common that they can induce predictable changes in the implicit and explicit sense of a bodily self. Yet, the question of what components of the bodily self are really altered during such illusions and how the various measures relate to them is still under debate. Longo et al. (2008) used a psychometric analysis of an extended questionnaire presented after the induction of the rubber hand illusion to identify three components of the illusion: (1) ownership, i.e., the perception of the rubber hand as part of oneself; (2) location, i.e., the localization of one's own hand or of touch applied to one's own hand in the position of the rubber hand; and (3) sense of agency, i.e., the experience of control over the rubber hand. These different components seem also to be reflected in differential neural activity as revealed by recent functional neuroimaging studies.[5]
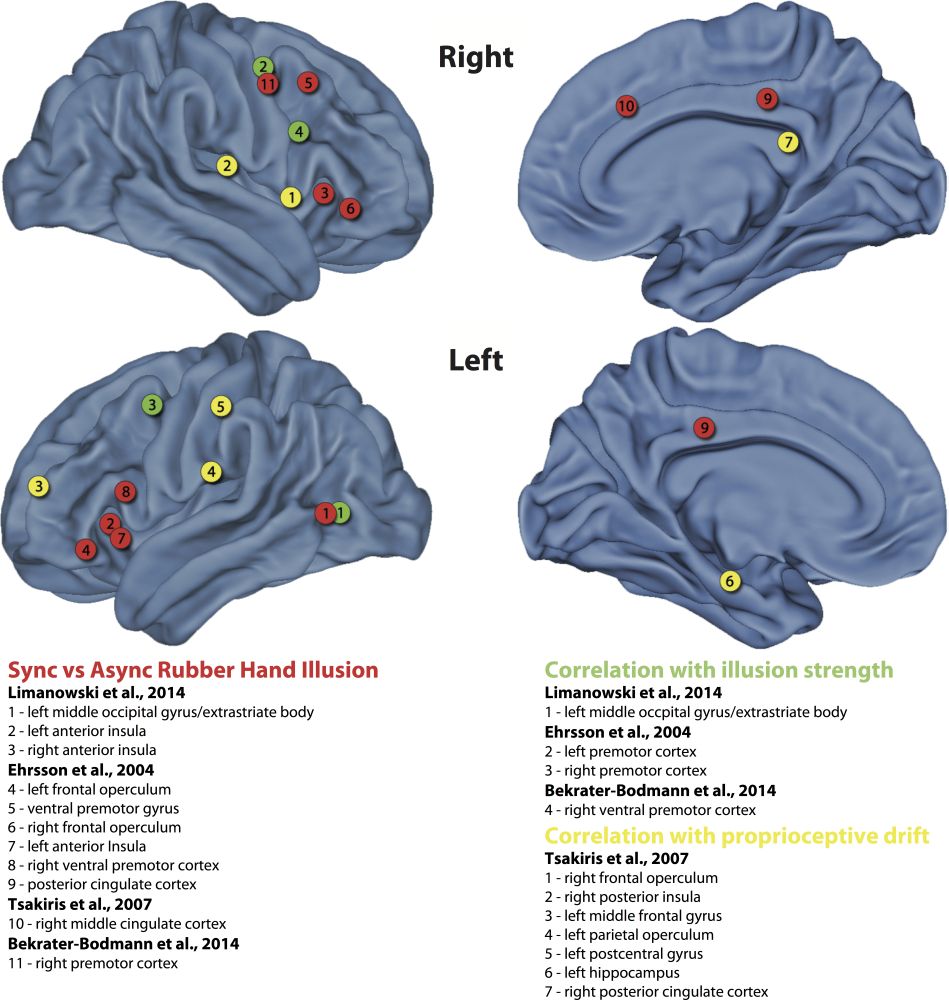 Figure 1: An overview of brain imaging studies of the rubber hand illusion (Bekrater-Bodmann et al.2014; Ehrsson et al.2004; Limanowski et al.2014; Tsakiris et al.2006). Red circles indicate significant brain activation in the comparison of synchronous visuo-tactile stimulation (illusion condition) to the control asynchronous visuo-tactile stimulation. Green circles indicate brain areas where the hemodynamic response correlates with the strength of the rubber hand illusion. Yellow circles indicate areas that significantly correlate with the proprioceptive drift. For the generation of the figure, MNI coordinates were extracted from the original studies and mapped onto a template with caret (http://www.nitrc.org/projects/caret/ (van Essen et al. 2001)).
Figure 1: An overview of brain imaging studies of the rubber hand illusion (Bekrater-Bodmann et al.2014; Ehrsson et al.2004; Limanowski et al.2014; Tsakiris et al.2006). Red circles indicate significant brain activation in the comparison of synchronous visuo-tactile stimulation (illusion condition) to the control asynchronous visuo-tactile stimulation. Green circles indicate brain areas where the hemodynamic response correlates with the strength of the rubber hand illusion. Yellow circles indicate areas that significantly correlate with the proprioceptive drift. For the generation of the figure, MNI coordinates were extracted from the original studies and mapped onto a template with caret (http://www.nitrc.org/projects/caret/ (van Essen et al. 2001)).
Figure 2 summarizes the main brain regions found to be involved in the rubber hand illusion during functional magnetic resonance imaging (fMRI) or positron emission tomography (PET) studies (Bekrater-Bodmann et al. 2014; Ehrsson et al. 2004; Limanowski et al. 2014; Tsakiris et al. 2006). The activation patterns depend on how the illusion was quantified. The pure contrast of the illusion condition (i.e., synchronous stroking) to the control condition reveals a network including the insular, cingulate, premotor, and lateral occipital (extrastriate body area) cortex. Areas in which haemodynamic responses correlate with the strength of illusory ownership include the premotor cortex and extrastriate body area, whereas illusory mis-localization of the physical hand (referred to as “proprioceptive drift”) correlates particularly with responses in the right posterior insula, right frontal operculum, and left middle frontal gyrus (see figure 1 for the detailed list). The fact that different brain regions are involved in illusory ownership and mis-localization of the physical hand provides further evidence for distinct sub-components underlying the bodily self.
2.1.2 Full-body illusions
Several authors claimed that research on body part illusions is unable to provide insight into the mechanisms of global aspects of the bodily self, such as self-identification with a body as a whole, self-location in space, and first-person perspective (e.g., Blanke & Metzinger 2009; Blanke 2012; Lenggenhager et al. 2007). Thus, empirical studies have more recently adapted the rubber hand illusion paradigm to a full-body illusion paradigm where the whole body (instead of just a body part) is seen using video-based techniques and virtual reality.
Two main versions of multisensory illusions targeting more global aspects of the self have been used (but see also Ehrsson 2007), one in which the participants saw the back-view of their own body (or a fake body) in front of them as if it were seen from a third-person perspective (full-body illusion [see figure 2, orange frame]; Lenggenhager et al. 2007) and one in which a fake body was seen from a first-person perspective (body swap illusion [see figure 2 green frame; Petkova & Ehrsson 2008]). In both versions of the illusion, synchronous visuo-tactile stroking of the fake and the real body increased self-identification (i.e., full-body ownership)[6] with a virtual or fake body as compared to asynchronous stroking. Importantly, it has been argued that only the former is associated with a change in self-location[7] (Aspell et al. 2009; Lenggenhager et al. 2007; Lenggenhager et al. 2009) and in some cases with a change in the direction of the first-person visuo-spatial perspective (Ionta et al. 2011; Pfeiffer et al. 2013).
A recent psychometric approach identified three components of the bodily self in a full-body illusion set up: bodily self-identification, space-related self-perception, which is closely linked to the feeling of presence in a virtual environment (see section 4.5.1.3), and agency (Dobricki & de la Rosa 2013). Again, these sub-components seem to rely on different brain mechanisms. Figure 2 contrasts two recent brain imaging studies using full-body illusions (see Serino et al. 2013, for a more thorough comparison). While self-identification with a fake body seen from a first-person perspective is associated with activity in premotor areas (Petkova et al. 2011), changes in self-location and visuo-spatial perspective are associated with activity in the temporo-parietal junction (TPJ) (Ionta et al. 2011). The TPJ is a region located close to the parieto-insular vestibular cortex (see section 3.2.3), suggesting that the vestibular cortex might play a role in the experienced self-location and visuo-spatial perspective, as we will elaborate on in the following sections.
2.2 Agency
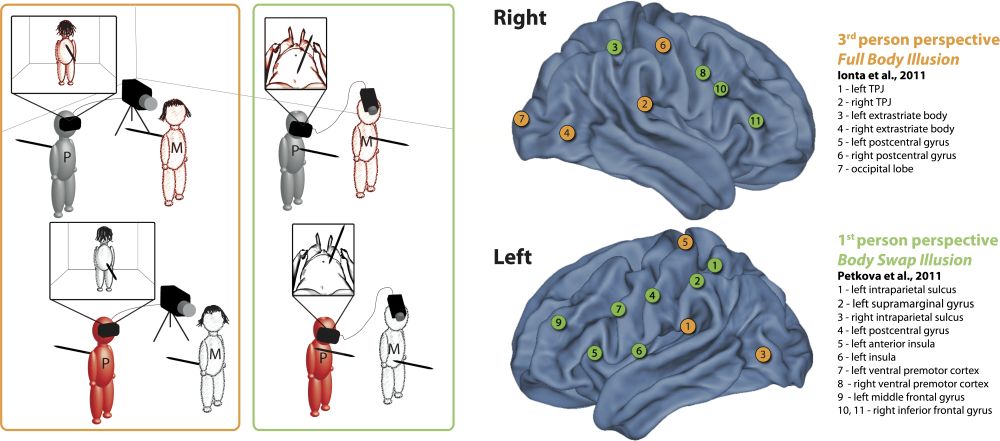 Figure 2: A comparison of brain activity associated with two illusions targeting the manipulation of more global aspects of the bodily self, i.e., the full body illusion (Lenggenhager et al.2007, setup in orange frame) and the body swap illusion (Petkova & Ehrsson 2008, setup in green frame). In both variants of the illusion, synchronous stroking of one’s own body and the seen mannequin led to self-identification with the latter (locus of self-identification is indicated in red colour). Two recent fMRI studies using either the full body illusion (Ionta et al. 2011 in orange circles) or the body swap illusion (Petkova et al. 2011, in green circles) are compared and plotted. Only areas significantly more activated during synchronous visuo-tactile stimulation (illusion condition), as compared to control conditions, are shown. For the generation of the figure, MNI coordinates were extracted from the original studies and mapped onto a template with caret (http://www.nitrc.org/projects/caret/). Adapted from Serino et al. 2013, Figure 2.
Figure 2: A comparison of brain activity associated with two illusions targeting the manipulation of more global aspects of the bodily self, i.e., the full body illusion (Lenggenhager et al.2007, setup in orange frame) and the body swap illusion (Petkova & Ehrsson 2008, setup in green frame). In both variants of the illusion, synchronous stroking of one’s own body and the seen mannequin led to self-identification with the latter (locus of self-identification is indicated in red colour). Two recent fMRI studies using either the full body illusion (Ionta et al. 2011 in orange circles) or the body swap illusion (Petkova et al. 2011, in green circles) are compared and plotted. Only areas significantly more activated during synchronous visuo-tactile stimulation (illusion condition), as compared to control conditions, are shown. For the generation of the figure, MNI coordinates were extracted from the original studies and mapped onto a template with caret (http://www.nitrc.org/projects/caret/). Adapted from Serino et al. 2013, Figure 2.
Agency, the feeling that one is initiating, executing, and controlling one's own volitional actions, has been described as another key aspect of the bodily self and self-other discrimination (Gallagher 2000; Jeannerod 2006; Tsakiris et al. 2007). Experimental investigations of the sense of agency started in the 1960s with a study by Nielsen (1963) In this seminal study, as well as in follow-up studies, a spatial or a temporal bias was introduced between a physical action (e.g., reaching movement toward a target) and the visual feedback from this action (Farrer et al. 2003b; Fourneret & Jeannerod 1998). These studies measured the degree of discrepancy for which the movement is still self-attributed. Theories of the sense of agency have mostly been based on a “forward model,” which has been defined in a predictive coding framework (Friston 2012). The forward model uses the principle of the efference motor copy, which is a copy from the motor commands predicting the sensory consequences of an action. Such efference copies allow the brain to distinguish self-generated actions from externally generated actions (Wolpert & Miall 1996). This idea is supported by a large body of empirical evidence showing that the sense of agency increases with increasing congruence of predicted and actual sensory input (e.g., Farrer et al. 2003a; Fourneret et al. 2001). Neurophysiological and brain imaging studies showed a reduction of activation in sensory areas in response to self-generated, as compared to externally generated, movements (e.g., Gentsch & Schütz-Bosbach 2011). As well as suppression of activity in specific sensory areas, agency has also been linked to activity in a large network including the ventral premotor cortex, supplementary motor area, cerebellum, dorsolateral prefrontal cortex, posterior parietal cortex, posterior superior temporal sulcus, angular gyrus, and the insula (David et al. 2006; Farrer et al. 2008; Farrer et al. 2003a).
While studies on agency have almost exclusively investigated agency for arm and hand movements, a recent study has addressed “full-body agency” during locomotion using full-body tracking and virtual reality (Kannape et al. 2010). As the vestibular system is importantly involved in locomotion, we will argue for a strong implication of the vestibular system in full-body agency during locomotion (see section 4.4).