3 Direction selectivity in SAC dendrites: Beyond componential constitution
Starburst Amacrine Cells are axonless neurons found in the retina of mammals and numerous non-mammalian species. Their morphology is planar, with multiple dendrites arrayed, as the name suggests, in a starburst pattern around the cell body (Figure 1).
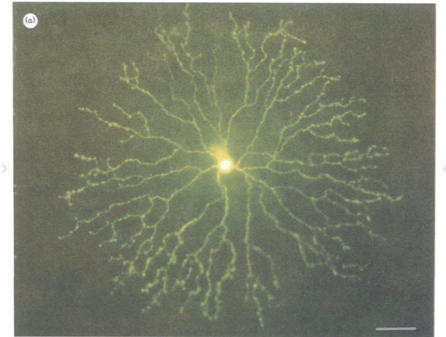 Figure 1: Micrograph of a Starburst Amacrine Cell. Calibration bar 50 µm. Reprinted from Tauchi & Masland (1984).
Figure 1: Micrograph of a Starburst Amacrine Cell. Calibration bar 50 µm. Reprinted from Tauchi & Masland (1984).
SACs form dense, highly overlapping, co-fasciculating layers in the “on” and “off” levels of the inner synaptic layer of the retina, nestled physically and functionally between bipolar cells and direction-selective ganglion cells. Among the most numerous neural cells found in the mammalian retina, they represent a large proportion of the total neural volume in the eye; in the rabbit retina, for example, as much as six meters of SAC dendrites occupy each square millimeter of retinal surface—higher coverage than any other retinal cell by an order of magnitude (Masland 2005; Tauchi & Masland 1984; see Figure 2).
 Figure 2: Depiction of the SAC network in peripheral retina. Calibration bar 50µm. Reprinted from Tauchi & Masland (1984).
Figure 2: Depiction of the SAC network in peripheral retina. Calibration bar 50µm. Reprinted from Tauchi & Masland (1984).
SACs are interesting for multiple reasons. Despite lacking axons, they synthesize and release both excitatory and inhibitory neurotransmitters (ACh (acetylcholine) and GABA (-Aminobutyric acid)) from the distal regions of their dendrites. Both the role and relative proportion of excitatory and inhibitory synaptic connections change over time. Cholinergic synaptic connections between neighboring SACs disappear over development, and GABAergic connections between SACs begin as excitatory but later become inhibitory. However, excitatory cholinergic synapses between SACs and ganglion cells remain (Masland 2005).
Functionally, SACs play an important role in motion detection, and are part of the overall network for multiple uses including optokinetic eye movement and motion perception (Yoshida et al. 2001). In fact, each dendrite of the SAC acts independently of the others, and signals the presence of stimuli moving centrifugally, that is, from the cell body out in the direction of the signaling dendrite (Euler et al. 2002; see Figure 3). Put differently, each SAC dendrite is a directionally selective spatial sub-part of the overall cell, and this is the functional property that will interest us here. As with so much in the neurosciences, the mechanism that explains this function is complex and not fully understood. It is, however, possible to offer a sketch of it.
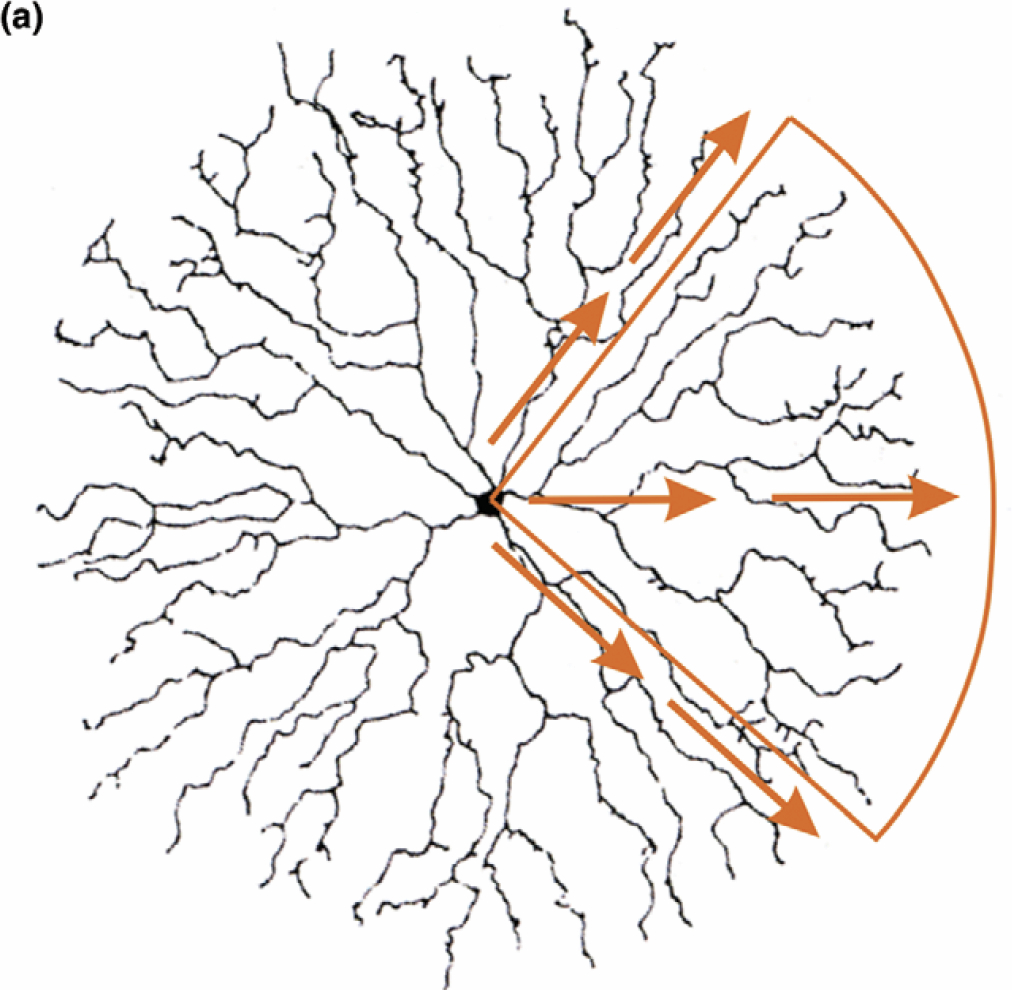 Figure 3: Depiction of direction selectivity in SAC dendrites. Reprinted from Masland (2005).
Figure 3: Depiction of direction selectivity in SAC dendrites. Reprinted from Masland (2005).
As mentioned above, SACs lie between bipolar cells and direction-selective ganglion cells. Bipolar cells thus mediate the initial stimulus such that a moving light causes them to fire in turn as the stimulus moves across the retina. The bipolar cells make excitatory synapses onto the SAC dendrites.[5] With these basic anatomical facts in view, we can turn to describing three different aspects of the overall mechanism for direction selectivity: wiring specificity between bipolar cells and the SAC dendrites; lateral inhibition between neighboring SACs; and active elements in the dendrites themselves.
First, the axonal projections of bipolar cells largely preserve the topography of their inputs, such that neighboring axons come from cells with neighboring inputs, and make neighboring synapses onto post-synaptic cells. What this arrangement means for SACs is that neighboring synapses on the dendrite are likely to come from neighboring bipolar cells, so that when a moving stimulus activates one cell, and then another immediately to its left (say), this will tend to activate a given synapse, and then another immediately to its left. Thus, in the case where such a stimulus moves along the direction of a dendritic process, the successive excitatory inputs to that dendrite will tend to reinforce (Demb 2007; Lee & Zhou 2006). This is an important part of the overall mechanism, but is not sufficient by itself to produce the observed directional selectivity, as these inputs would tend to reinforce even during centripetal motion, although this would result in a weaker response at the distal process of the dendrite (Hausselt et al. 2007).
Another important part of the mechanism for directional selectivity involves mutual inhibition between neighboring SACs (Figure 5). As a stimulus moves so as to stimulate the centrifugal dendrite of SAC1 (in Figure 5A), reinforcing inputs will cause the release of GABA onto the centripetal dendrite of SAC0, such that even when the light stimuli begins to excite the centripetal dendrite of SAC0, the leading inhibition dominates the signal. Similarly, as the stimulus moves to the centrifugal dendrite of SAC0, the successive excitatory inputs from the bipolar cells reinforce, and any inhibitory inputs from the neighboring SAC2 come too late. Moreover, SAC0 will largely inhibit SAC2’s response (Figure 5B; Lee & Zhou 2006). An important element of this mechanism involves the relative time-course of ACh and GABA: ACh response from the bipolar cells ramps up and decays fairly quickly, while GABA response is relatively delayed and prolonged (Demb 2007). This temporal asymmetry helps ensure that when inhibition leads it dominates, and vice-versa. The distance between SACs also plays a role. The likelihood of synaptic connections between the distal portion of the dendrites of two SACs—where inhibitory connections are most effective—depends on the distance between the cell bodies. Cells that are very close together or very far apart will thus not mutually inhibit one another (Figure 5C).
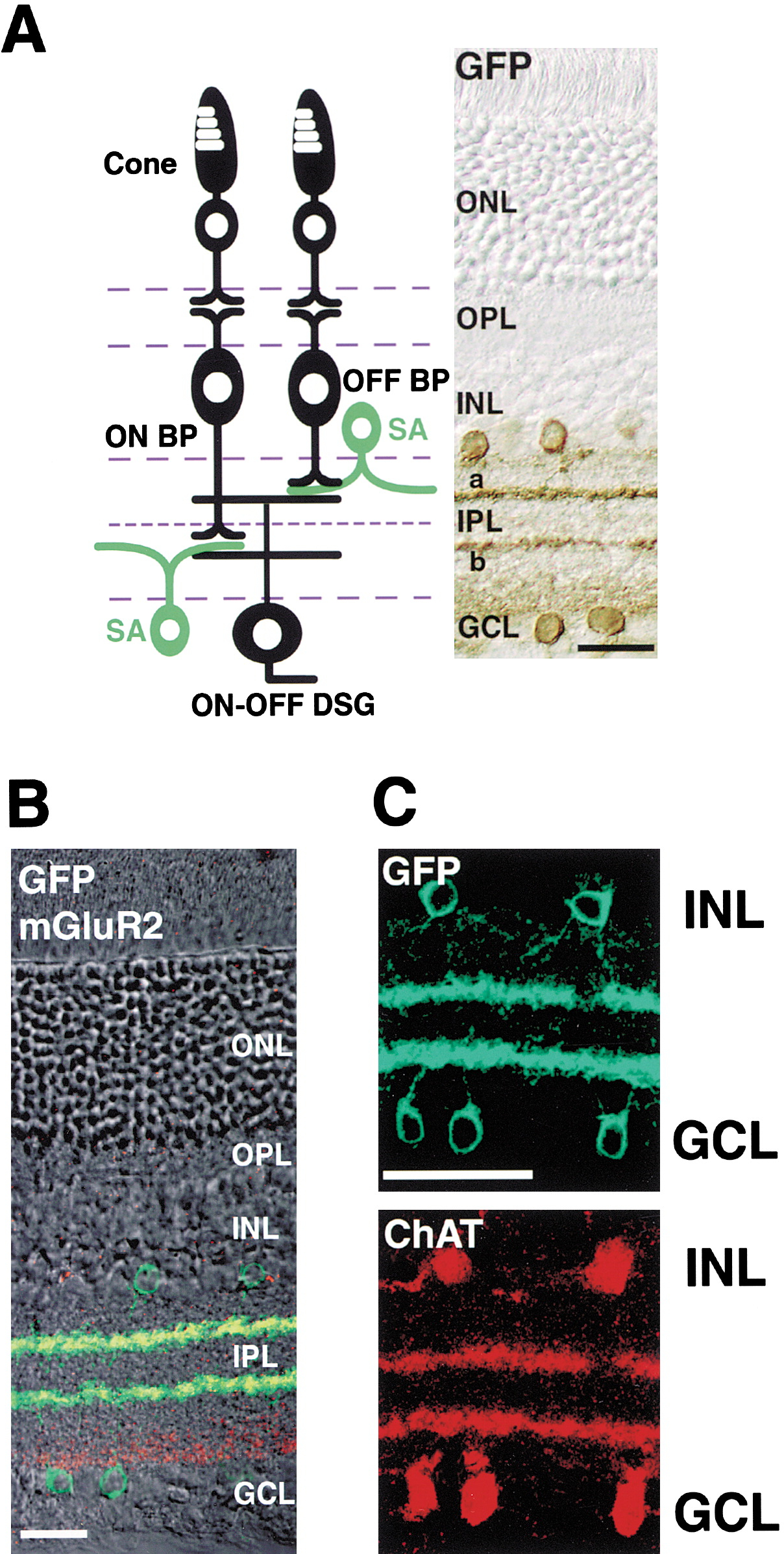 Figure 4: Schematic representation of the layered structure and synaptic relationships between bipolar cells and SACs. Reprinted from Yoshida et al. (2001).
Figure 4: Schematic representation of the layered structure and synaptic relationships between bipolar cells and SACs. Reprinted from Yoshida et al. (2001).
Finally, direction selectivity depends upon properties of the dendrite itself. The dendrites are electrically isolated from one another, as a result of both overall cell morphology and the low impedance of the cell body. The uneven distribution of synaptic inputs and outputs also contributes: excitatory inputs from the bipolar cells are distributed along the length of the dendrite, but synaptic outputs are confined to the distal ends (as implied by the two aspects of the overall mechanism described above). A third, active aspect of the local dendritic portion of the mechanism appears to involve voltage-gated calcium channels. These channels lead to amplification of the ACh response beyond what the passive reinforcement caused by successive synaptic transmission from bipolar cells can account for (Hausselt et al. 2007).
All of these elements combine to produce the direction selectivity of the SAC dendrite. Bipolar cells successively synapse onto the dendritic process, resulting in passive reinforcement of excitatory input that preferentially promotes neurotransmitter release in response to motion in the centrifugal direction. Surrounding SACs selectively inhibit centripetal excitation, as a result of the different temporal activation profiles of GABA and ACh; the asymmetric distribution of input and output synapses; and the relative spatial placement of the SACs. And voltage-gated calcium channels in the dendrite actively amplify the centrifugal signal. Although this sketch leaves out many of the known details, and there remain many details still to be worked out, I believe it is sufficient to warrant the conclusion that this is (a) an instance of mechanistic explanation that (b) does not have the level-restricted character of the (canonical) mechanistic explanations laid out above. I spell out the reasons for this conclusion in the next section.