6 Parts and wholes
A distinct and indirectly related application of the levels metaphor in the neighborhood of levels of realization invites a different kind of equivocation, this time concerning the existence of higher-level powers. This application involves not a whole-whole relationship but rather the relationship between the behavior or property of a whole and the behaviors or properties of one of its parts. The behavior of the whole does not (except in special cases) supervene on the operation of the individual parts. The grain of sand contributes to the mass of the sand pile. The kidney contributes to the capacity of creatures to maintain plasma osmolality. In each case, the property of the whole (the mass of the pile, the regulation of plasma osmolality) might differ even when the contribution of these singular parts remains the same. In part-whole levels, as opposed to levels of realization, the relationship is between parts and wholes, not between wholes and the corresponding organized collections of entities, properties, and activities. In this case, eliminativism about levels is a nonstarter (whatever its metaphysical credentials); it is impossible to imagine neuroscience, biology generally, or indeed most special sciences without the idea that things have parts.
In applying the levels metaphor to this part-whole relation, one emphasizes the relations question over the placement question.[11] In levels of size, things are at a given level because they are similar in size. Levels, thus conceived, are monolithic: they reach across all of nature, embracing everything within a given size range. Oppenheim and Putnam’s layered model of the world might be read as similarly monolithic. Wimsatt’s tree diagram breaks with this monolithic view precisely because it emphasizes compositional relationships: different branching levels are required because different kinds of whole (cosmological objects, human societies) are composed in different ways. If one centers part-whole thinking in one’s application of the levels metaphor, then the metaphor carries no particularly useful answer to the placement question. One can offer only a necessary condition: two things are at the same level only if they are not related as part to whole. Given that most things are not related to one another as part to whole, the resulting idea of being “at” a part-whole level has little or no conceptual significance.
Levels of parts and wholes lack many of the features in Wimsatt’s prototype of levels. Many of the features in that prototype appear to derive from the monolithic conception shown in Figure 3: causes, forces, and laws are most plausibly thought to cluster together on the assumption that size is relevant to which forces can act, that causal relations are expressions of forces acting, and that laws govern these interactions. But if one places the part-whole relation in the center of one’s metaphor, then there is no reason to embrace an empirical association between being at a given level and having a proprietary set of causal interactions for that level. Levels of parts and wholes must also be correlated with size differences because parts can be no larger than the wholes they compose. But the size differences between levels of parts and wholes are an accidental consequence of the part-whole relationship itself, not part of defining what it is for things to be at different part-whole levels.
In the following subsections, my goal is to sketch some contours of the relevant notion of part and whole. I start with classical mereology only to make the point that this apparatus was not constructed with an eye to developing a descriptively adequate account of the levels described by science. The more we learn about the limits of these classical models for our present purposes, the more we place constraints on the relevant notion of levels that, as I and others have argued, is central to the explanatory structure of neuroscience and the special sciences generally: levels of mechanisms (Bechtel 1988; Bechtel & Richardson 1993; Craver 2001; Machamer et al. 2000).
6.1 Types and tokens of parts and wholes
The Gibson SG has two humbucker pickups. My Gibson SG has two humbucker pickups. Not all Gibson SG’s have two humbucker pickups. But that’s the factory model, the central exemplar or prototype against which variations are evaluated as more or less “typical.” Likewise, when we talk about the human brain or the frog kidney, we are talking about types: the human type of brain, the frog type of kidney. And we talk also about the parts these types of things typically have.
The monolithic, layer-cake image in the Oppenheim-Putnam hierarchy is a mereology of types: societies are formed of organisms, organisms of cells, cells of molecules, and so on. Wimsatt’s tree also represents relationships between types. Crystals are made of crystalline solids, which are made of inorganic molecules, which are made of atoms. It is true, of course, that all cells are made of molecules and that all organs are made of cells. But is not true that all cells are at a higher level than molecules generally.
It seems natural and harmless enough to treat the part-whole relations among types as generalizations over relations between particular wholes and particular parts. When one says, “The human brain has two hemispheres and a corpus callosum,” one asserts that having these parts is typical of human brains. One is warranted on the basis of such a claim (though not always correct) in asserting of a particular human brain that it has two hemispheres. That is, the relation among part and whole types derives from a more primitive, token relationship between particular parts and wholes. Type-level claims about part-whole relations assert that such part-whole relations regularly or typically hold in the individuals in the relevant reference class.
One of the many useful insights contained implicitly in the branching structure of Wimsatt’s tree diagram (Figure 1) is that different types of wholes are made up of different types of parts and are naturally decomposed into different levels. Both the human brain and the frog kidney are organs, and both are made of cells, but the cells in each are different, and these cells are organized differently into higher-level components. If we look within the human brain, we find that different brain regions are composed of altogether different components and exhibit more or less proprietary organization. Broadman mapped the brain by studying these differences in cytoarchitecture from one brain region to the next. The receptive field organization of the visual cortex has, to my knowledge, no companion in the organization of the amygdala or of the mammillary bodies. Different types of brain regions/systems are made up of different types of components: they have different part-whole levels.[12]
When we say that objects or processes of one type are parts of objects or processes of another type, we are asserting that ceteris paribus token objects of the one type are composed of token objects of the second type. Indeed, standard attempts to define the part-whole relationship with logical rigor are expressed in terms of relationships among token individuals (Varzi 2014).[13]
6.2 Mereology
Although Oppenheim and Putnam describe the layer-cake structure of science in terms of different types of objects, the mereological structure they use to support this picture is expressed in terms of tokens. Classical mereology provides a very general and content-neutral account of the part relation. It can be used equally well to express the sense in which the word apple has the letter “a” as a part, in which courage has judgment as a part, and in which apple pie has cinnamon as a part. Common axioms associated with classical mereology, including the mereology adopted by Oppenheim and Putnam (Rescher & Oppenheim 1955), include:
1) Reflexivity: Every object is a part of itself.
2) Transitivity: Every part of a part of an object is part of the object.
3) Extensionality: An object is completely determined by the set of its parts; i.e., for objects to be identical, it suffices that they have all and only the same parts.
4) Summation: Any pair of objects (x, y), is itself an object, z, which is their sum.
I list these constraints only to illustrate that classical mereology will take us only so far in the effort to characterize part-whole levels. This is a formal theory, abstracted entirely from the concerns of practicing scientists. This means that there are constraints in the scientific conception of parts and wholes that classical mereology need not honor. First, the levels that Churchland and Wimsatt describe are space-, structure-, and time-involving in ways that classical mereology need not be. The set of integers is part of the set of real numbers, and “Consider the Lobster” is part of Wallace’s corpus, but not in the same way that the glutamate receptor is part of the chemical synapse. The glutamate receptor takes up part of the space occupied by the synapse as a whole. Its opening is part of the extended process by which neurons communicate. None of this is expressed or intended to be expressed in the generic part-whole relation of classical mereology.[14]
Second, although reflexivity is involved in certain theoretical applications of mereology, it has no application in thinking about such space-involving levels. It does no justice to the biological concept to assert that every hippocampus is a part of itself. If levels are defined as a relationship between a part and a whole, and everything is a part of itself, then everything is at both higher and lower levels than itself. The parts surely must be proper parts.[15]
Third, it has been noted that the transitivity axiom often fails to apply to functional parts of the sort that populate physiological and biological theories (Varzi 2014). Eric is part of the championship pool team, and Eric’s locks are part of Eric, but his locks are not part of the team. However, if one requires of a part (entity, property, or activity) that it must be relevant to the property or behavior of the whole, then one can retain the transitivity of this relation, at least in many contexts. If we ask not about Eric, but about the motion of his arm as he wields his cue, then his locks are clearly not relevant while the muscles gliding his arm steadily forward are relevant. So too are, in some sense, the molecules transmitted across the neuromuscular junction during his backstroke. When what counts as a part is filtered in each iteration by explanatory relevance relations (they are not mere spatial or temporal parts but working parts—parts that are involved in, contribute to, or make a difference to the property or activity of the whole), then the relationship is, in fact, transitive. The appearance of a failure of transitivity in functional systems trades, it seems, on failing to relativize the decomposition into parts by a highest-level target (explanandum) phenomenon; not all the spatiotemporal parts of an object or process are relevant to everything it does. It is only relative to a highest-level activity or property of the hierarchy under consideration that the lower level parts are visible as components—as working parts in the mechanism. If we think not about the team, Eric, and his locks, but rather about the victory, the shot, and the muscular contraction, matters seem different.[16] The contraction contributes to the shot, which contributes to the victory. The locks will not appear in this hierarchy, but the relevant parts will.[17]
The extensionality theorem holds that no two distinct objects share all and only their proper parts. A hippocampus and a bust of the Dalai Lama formed out of the same pyramidal cells, granule cells, etc. that compose the hippocampus are, according to classical mereology, one and the same object. But in biological systems, the organization of components is often (perhaps always) relevant to the properties and activities of the whole. Again, parts appear as parts only relative to a decomposition framed by reference to some highest-level property or activity. This is Kauffman’s point, enshrined in Glennan’s law: a mechanism is always a mechanism of a given phenomenon (Kauffman 1971; Glennan 1996). Thus Kauffman:
A view of what the system is doing sets the explanandum and also supplies criteria by which to decide whether or not a proposed portion of the system with some of its causal consequences is to count as a part and process of the system. Specifically, a proposed part will count as a part of the system if it, together with some of its causal consequences, will fit together with the other proposed parts and processes to cause the system to behave as described. (1971, p. 260)
The more general point is that there is an application of the levels metaphor that is not merely a part-whole relationship as specified in classical mereology, but one in which the parts are relevant (explanatorily and constitutively) to some property or activity of the whole.
One can make a similar point with respect to the summation axiom. This theorem allows one to form arbitrarily many gerrymandered wholes out of disparate and unconnected parts with no spatial, temporal, causal, or functional unity. Lewis (1991) calls this “unrestricted composition”: whenever there are some things, there is also a fusion of those things. The Yankees’s starting rotation and the now disparate parts of my mother’s old Chevy Vega together form a whole. This way of thinking about parts and wholes has little or no application in biology because such gerrymandered wholes don’t do anything interesting (though such wholes will have aggregate properties of the sort discussed below). The whole in such gerrymandered collections typically doesn’t play any explanatory role. And what goes for wholes goes for parts as well. According to this classical picture, it is perfectly legitimate to claim that my dog, Spike, has four parts: a front quarter, a hindquarter, and two midsections of approximately equal length. There is nothing to prevent this way of talking; but the parts revealed in this decomposition do not cut Spike at his joints. The biological decomposition finds joints at causal interfaces, and identifies parts with more or less isolable (nearly decomposable) subsystems (Simon 1962) that contribute to the behavior of the whole.
In short, many of the ideas central to classical mereology must be amended or restricted if they are to apply to the part-whole levels distinctive of biology, neuroscience, and the special sciences generally. At least some of the work can be done by restricting the part-whole relation by a relevance condition on biological parthood: all the lower-level properties, activities, and organizational features of the parts are relevant to—contribute to— the property or activity of the whole.
6.3 Levels of organization: Aggregates and mechanisms
So let us focus on an application of the levels metaphor that is a part-whole relation and a (constitutive) relevance relation. I will not dwell here on the appropriate notion of relevance (see Craver 2007, see also Harinen 2014). For now, we can work with the idea that each part in such a hierarchy (in addition to being spatially and/or temporally contained within the whole) plays a necessary but insufficient role within a collection of parts that are jointly sufficient (but possibly redundant) for a given explanandum phenomenon (Couch 2011). That is, relevant parts might usefully be thought of as constitutive INUS (insufficient, but necessary part of an unnecessary but sufficient condition) for the behavior of the mechanism as a whole (Mackie 1973).[18]
Again following Wimsatt (1997), we can distinguish two ways that spatiotemporal parts contribute to a property or activity of a whole: aggregation and organization. An aggregate property is literally a sum of the properties or activities of the parts. The current flowing through an ion channel, for example, is a sum of the charges carried by individual ions. The concentration of a volume of a fluid is a sum of the number of particles in that unit volume. Aggregative properties change linearly with the addition and removal of parts. And aggregative properties do not change as the parts are intersubstituted with one another. Some properties of the hippocampus, such as its mass, remain the same when the cellular constituents of the hippocampus are reorganized to represent His Holiness. Other properties of the hippocampus, such as its information processing capacities, are destroyed. For truly aggregative properties, spatial, temporal, and causal organization among the components is irrelevant.
Aggregates are rare. The masses of the individual grains in a sand pile do, in fact, depend on the spatial distribution of the other grains (if one takes relativity seriously). What is presumed to be a homogeneous concentration of a liquid can in fact have local concentration differences depending on how the ions are organized in different parts of the fluid. In the case of non-aggregates, the activity or property of the whole is not a simple sum of the properties of the individuals. Adding or removing parts (e.g., the human heart) can lead to dramatic changes in how the system (e.g., the body) works. And rearranging the parts and their activities in space and time can eliminate the explanandum phenomenon entirely (as would happen if one randomly swapped parts of the circulatory system for one another). This is all true because spatial, temporal, and causal organization are relevant to (make a difference to, partly constitute) the property of the whole.
I use the term “mechanism” permissively to describe non-aggregative compositional systems in which the parts interact and collectively realize the behavior or property of the whole. Mechanisms are by definition more than the sums of their parts: they have properties their parts do not have, and they engage in activities that their parts cannot accomplish on their own.
Most mechanisms with which I am familiar involve myriad part-whole relations, some of which are more aggregative in nature, and some of which are less so. Many things brains do, for example, involve the flux of ions across a membrane, which flux is closer to the aggregative than the mechanistic end of the organizational spectrum. Other things brains do (such as the developing grid cells in the entorhinal cortex) require precisely organized relations among the activities of cells in and around the entorhinal cortex. This organizational spectrum from aggregate to mechanism covers all the relations that go into levels of organization, the superordinate class.[19]
In levels of mechanisms, the relata are some activity or property of a mechanism as a whole,[20] and the activities, properties, or organizational features of its components (its relevant parts and organization). Some component, X’s φ-ing, is at a lower mechanistic level than S’s ψ-ing if and only if X’s φ-ing is a component in S’s ψ-ing, that is, if and only if X’s φ-ing is a relevant spatiotemporal part of S’s ψ-ing. In levels of mechanisms (as opposed to aggregates) lower-level components are organized together to make up some behavior or property of the whole; in aggregates, the properties of the parts are summed.
Levels of mechanisms are represented in Figure 4. At the top is the activity of some mechanism as a whole (S’s ψ-ing). S’s ψ-ing is a behaving mechanism. Although one can speak of the mechanism and its activity separately (as when a mechanism stands inactive but ready to act), such separation in thought is artificial. Even the static mechanism is defined and sub-divided by reference to what it does. ψ is the topping-off activity of the mechanism for which all lower-level components are relevant. It can be idealized as an input-ouput relation, though this is an impoverished way of understanding phenomena (see Craver 2007). One level down are the activities and components, the X’s φ-ing, which compose and are organized together to constitute S’s ψ-ing.[21] Below that is another iteration of levels: the ρ-ings of Ps organized such that one of the Xs φs as it does. By organization, I mean that the parts have spatial (e.g., location, size, shape, and motion), temporal (e.g., order, rate, and duration), and active (e.g., feedback or other motifs of organization; see Levy & Bechtel 2014) relations with one another by which they work together to do something they cannot do on their own. As noted above, the relationship between levels is a part-whole relationship filtered further by constitutive relevance (Craver 2005; Harinen forthcoming). In levels of mechanisms, parts are made into higher-level components by being organized spatially, temporally, and actively into something. In more aggregate compositional relationships, they are summed into higher levels.
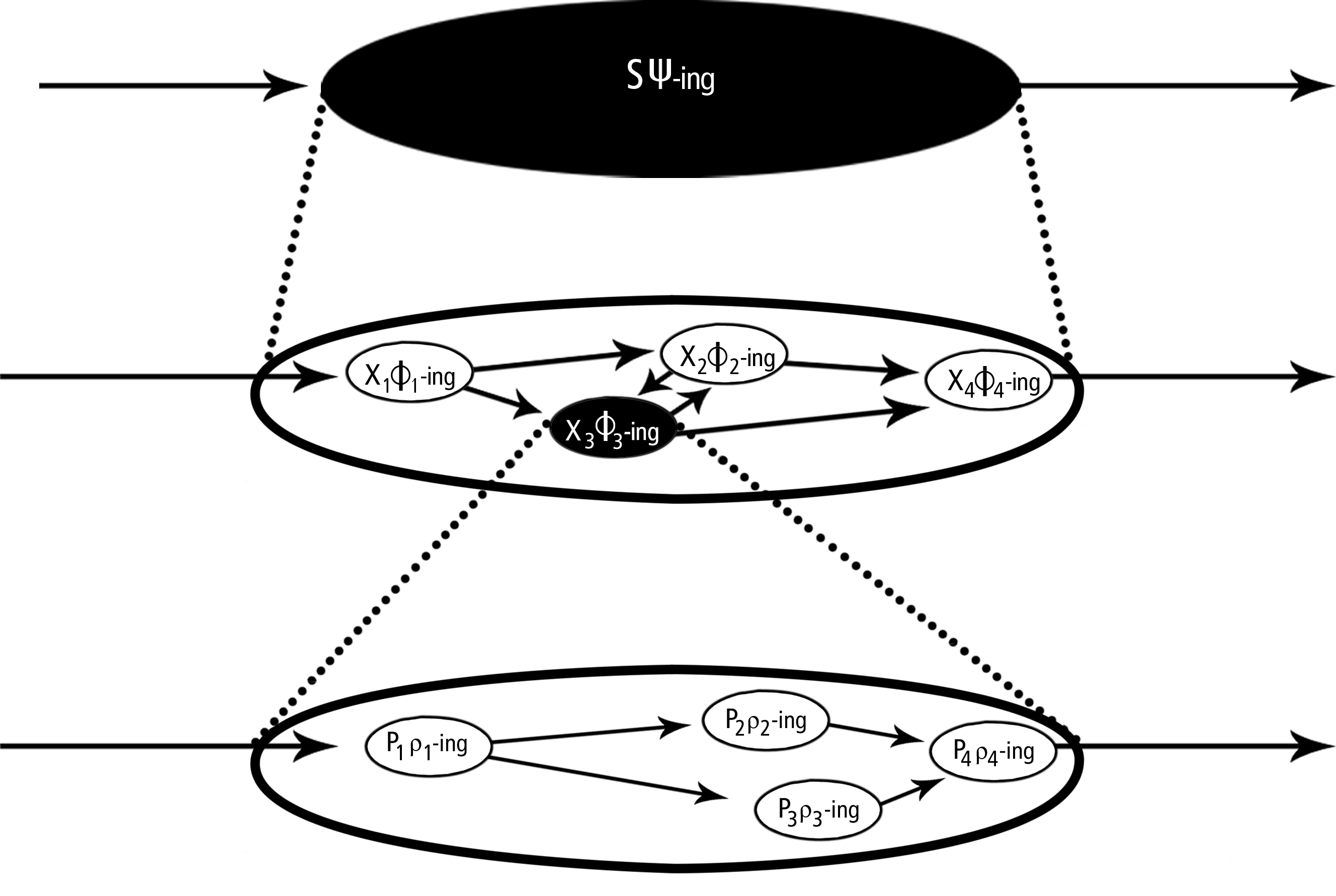 Figure 4: An abstract diagram of levels of mechanisms.
Figure 4: An abstract diagram of levels of mechanisms.
Contemporary theories of learning and memory provide a compelling example of levels of mechanisms (see Craver & Darden 2001; Craver 2002). The top level is a mechanism as a whole engaged in a spatial memory task, such as learning to run efficiently through a maze. One component in that mechanism, and so one level down in this description, is the hippocampus, a region of the brain thought to form a “map” of locations and orientations within the maze. The capacity of the hippocampus to acquire such an internal map of local spaces is thought to be explained, in part, by changes in synapses between pyramidal cells, specifically by a process known as Long-Term Potentiation (LTP). And it is now known that n-methyl d-aspartate (NMDA) receptors (n-methyl d-aspartate is a pharmacological agonist that binds these receptors preferentially), contribute to LTP. This story could continue downward, looking into aspects of protein chemistry and the structural changes thought to underlie channel functioning.
6.3.1 Levels of mechanisms are local
Levels of mechanisms are of entirely local significance. The levels in our example are defined by reference to a topping-off point, spatial memory, contribution to which determines whether or not a spatiotemporal part of the system is in fact relevant—whether it is a component in the mechanism for S’s ψ-ing. The hierarchy in Figure 4 and the levels of spatial memory as I have described them follow only a single (local) strand of embedding: from the behavior of the mechanism as a whole, to the behaviors of its components, on to the behaviors of one of these components, and so on.
Levels of mechanisms, like part-whole levels generally, are not monolithic divisions in the furniture of the world. Levels of mechanisms are defined only within a given part-whole hierarchy. There are different levels of mechanisms in the spatial memory system, in the circulatory system, in the osmoregulatory system, and in the visual system; the levels in each need not map onto one another. How many levels there are, and which levels are included, must be determined on a case-by-case basis by discovering which sorts of components are explanatorily relevant for a given phenomenon. Levels of mechanisms cannot be read off a menu of levels in advance.
If we apply the levels metaphor only locally, then it makes no sense to ask whether the hippocampus is at a higher or lower level than the nephra in the kidney. The nephra are not part of the hippocampus, and they are not relevant to the functioning of the hippocampus. Neither similarities of size nor similarities in kinds of part are definitive of levels of mechanisms. Rather, levels of mechanisms are defined relative to one another within a hierarchically organized mechanism.
The idea that levels of mechanisms retain some hint of the layer-cake model can sneak its way back into one’s application of the metaphor if one slides unknowingly between tokens and types of parts and wholes. Compare the following three sentences:
This pyramidal cell is at a lower level of mechanisms than this hippocampus.
Pyramidal cells are at a lower level of mechanisms than hippocampi.
Cells are at a lower level of mechanisms than organs.
Statement (a) expresses a mechanistic notion of levels: a particular pyramidal cell is a component of a particular hippocampal mechanism. This statement is true if the cell is a component in a mechanism for a given activity in which the hippocampus is engaged. It might be, for example, that a given pyramidal cell is a component in some hippocampal mechanisms but not others; if so, it is at a lower level to some hippocampal activities and not others.
Wimsatt describes the compositional relationship between levels as a relationship between types. He writes: “Intuitively, one thing is at a higher level than something else if things of the first type are composed of things of the second type” (Wimsatt 1976, p. 215). This is a departure from the idea of levels of mechanisms and one that threatens to reinstate something like the Oppenheim and Putnam hierarchy. Pyramidal cells are found outside the hippocampus, and those pyramidal cells are not parts in hippocampal mechanisms; they are not at a lower level of mechansitic organization. Likewise, both the hippocampus and the kidney are composed of cells; organs tend to be composed of cells. But the cells in the hippocampus are not at a lower mechanistic level than kidneys because they do not contribute to kidney function. The slide from sentences such as (a) to sentences such as (b) and (c) is a slide back toward the layer-cake model. Of course, scientists typically deal with types. But as I suggest above, this is a generalization over a relationship between tokens. The correct generalization is that the cells that compose hippocampuses are at a lower level than the hippocampuses they compose.
This is significant for two reasons. First, it helps to show that many objections to thinking about neuroscience and other special sciences in terms of levels simply do not apply to this restricted application of the metaphor. If one thinks, with Wimsatt, me, and probably Oppenheim and Putnam, that the Oppenheim and Putnam layer cake is an overly simplistic representation of the diverse ontological structures one finds in the special sciences—that things like occular dominance columns and synapses don’t readily fit that picture—one can nonetheless retain the idea that mechanisms are susceptible to multiple nested decompositions. These are different applications of the level metaphor. Secondly, and perhaps more importantly, the idea that levels are local significantly shifts the reductionist world-view for which Oppenheim and Putnam developed their ontology of levels. If one thinks of levels as levels of organization (as levels of mechanism and levels of aggregation), then it is inaccurate to think of reduction as involving relationships among theories developed to describe the items at a particular monolithic level. If reduction is simply a matter of explaining a higher-level phenomenon in terms of the organized activities of components, then reduction is still possible within a mechanistic world-picture, but it will be achieved not through grand reductions of overarching theories, but rather through piecemeal explanatory achievements for specific phenomena. Visions of the unity of science through interlevel reduction have to be revisioned not as grand unifications across the whole of science but rather as local explanatory successes. Such local explanations will, in fact, integrate findings from different sciences and bring different theoretical vocabularies into conversation with one another (see Craver 2005; Craver & Darden 2001), but it only deceptively resembles the layer cake that Oppenheim and Putnam sketched as a working hypothesis.[22]
6.3.2 Placement is weak and derivative in levels of mechanisms
One consequence of the mechanistic application of the levels metaphor is that there is no unique answer to the question of when two items are at the same mechanistic level. Only a partial answer is avialable: X’s φ-ing and S’s ψ-ing are at the same level of mechanisms only if X’s φ-ing and S’s ψ-ing are components in the same mechanism, X’s φ-ing is not a component in S’s ψ-ing, and S’s ψ-ing is not a component in X’s φ-ing.[23] Unlike size levels or levels defined in terms of the types of objects found at a given level, levels of mechanisms are defined fundamentally by the relations question: by the componency relationship between things at higher and lower levels. If two things are not related as part to whole, they are not at different levels, and so, if they are in the same mechanism, they are, in this very weak sense, at the same level. But this is just to say that sameness of level has no significance within this application of the metaphor.[24]
If one thinks of levels of organization as levels of aggregates and levels of mechanisms, then spatial containment and size relations between levels follow as an accidental consequence of the componency relation. The pyramidal cells are contained within the hippocampus, which is contained within the spatial memory system. The activities of these entities are also related as temporal part to whole: the binding of glutamate is a temporal component in the activity of the NMDA receptor. The objects at lower levels are smaller than (or at least no larger than) the whole, giving the hierarchy a derivative size ordering. Relations of size, rather than defining what it is for an item to be at a level (the placement question), are derivative from the more fundamental relationship between levels (the relations question): namely, the relationship between a mechanism and a component.
6.3.3 Emergence and levels of mechanisms
Mechanisms do things that their components taken individually cannot. This marks a sharp distinction between levels of mechanisms and levels of realization. Kim says this point is “obvious but important”:
This table has a mass of ten kilograms, and this property, that of having a mass of ten kilograms, represents a well-defined set of causal powers. But no micro-constituent of this table, none of its proper parts, has this property or the causal powers it represents. H2O molecules have causal powers that no oxygen and hydrogen atoms have. A neural assembly consisting of many thousands of neurons will have properties whose causal powers go beyond the causal powers of the properties of its constituent neurons, or subassemblies, and human beings have causal powers that none of our individual organs have. Clearly then macroproperties can, and in general do, have their own causal powers, powers that go beyond the causal powers of their micro-constituents. (Kim 1998, p. 85)
Through aggregation or organization, wholes have causal powers that their parts individually do not have. An activity at a higher level of mechanistic organization is quite literally more than the sum of its parts. It is not an aggregate. The addition and removal of parts leads to nonlinear changes in the behavior of the mechanism as a whole. It matters how the parts are organized; it is in virtue of their organization that they have properties that go beyond the properties of the individual parts (Wimsatt 1996). This feature of levels of mechanisms is so obvious, so prosaic, and so banal as to be hardly worth mentioning. No fancy complexity is required: two toothpicks stacked perpendicular to one another have the mechanistically emergent capacity to act as a lever or catapult; neither toothpick can do so on its own.
Of course, most mechanisms in biology are substantially more complicated than that. They have many more parts. Those parts interact with one another with bewildering complexity. Often they contain feedback relations that introduce nonlinear interactions into the operation of the mechanism itself. The mechanisms of LTP, for example, have yet to yield their secrets completely despite the dedicated attention of thousands of researchers over forty-odd years. A glance at any recent textbook is enough to convince one that LTP involves myriad intracellular reactions, protein synthesis, structural features of dendritic spines, changes to vesicular release, and retrograde transmission with nitric oxide. The mechanism involves so many parts and interactions that it would be useless, if not impossible, to represent them all in a visual diagram. Keeping track of how they all work together would require a very complicated computational model of some sort that has yet to be developed. As mechanisms get this complicated, we reach the limits of our ability to predict how the behavior of the whole will change as the parts change. Any change introduced to a part has so many ramifications that it is difficult or impossible for creatures like us to keep track of them all. This is an interesting fact about us and the limits of our cognitive and modeling prowess. But, ontologically, it is the same old banal fact about the importance of organization in mechanisms. We have added only that we have difficulty keeping track of it all.
Likewise, a common scientific complaint against reductionistic research programs in biology and neuroscience is that one can make only limited progress by studying the parts of mechanisms in isolation from one another.[25] We can study LTP in cells grown in a culture, and we can study hippocampal computations in a razor-thin hippocampal slice, and we can study spatial learning in highly contrived environmental settings such as a large pool filled with milky water (the Morris water maze). Such reductionist practices are absolutely essential to progress in the sciences. Nonetheless, one engaged in such practices must (and typically does) bear in mind that the behavior of the part when it is isolated for experimental purposes might be very different from the behavior the part exhibits when it is working in the context of a mechanism. Causal interactions with other parts of the mechanism and background conditions “in the wild” might lead to behaviors that would never be discovered in such simplified preparations. This is an extremely important point about reductionist research programs (Bechtel & Richardson 1993), and one might choose to describe this well-known difficulty with the language of emergence. But this is just to say that one cannot truly understand how a mechanism works until one understands how all its parts are organized together and working in the relevant conditions, and this we have already said repeatedly.
I emphasize the banality of these observations to stress that many of the things one wants to say about organization in biological systems can be said within the mechanistic application of the levels metaphor without introducing anything that is metaphysically novel or suspect. As the complexity of a mechanism increases, the epistemic challenges we face in discovering and modeling it increase as well, but this is of no significance for the ontic structures—the entities, activities, and organizational features that exist in the world.
Not so for spooky emergence. Spooky emergence is spooky precisely because it is commited to the existence of higher-level properties that have no explanation in terms of the parts, activities, and organizational features of the system in the relevant conditions. Levels of mechanisms are levels of ontic mechanistic explanation (Craver 2014): they are defined in terms of componency and constitutive explanatory relevance. If that explanatory relationship is severed, then the sense in which emergent properties are at a “higher level” must be altogether different than the compositional notion of levels in levels of mechanisms. If one imagines that atoms compose molecules, which are organized into cells, which are linked into networks from which mental properties spookily emerge, the first three steps are upward steps in a hierarchy of levels of mechanisms, but the last is not. The ability of organization to elicit novel causal powers (that is, nonaggregative behaviors and properties) is unmysterious both in scientific common sense and common sense proper (Van Gulick 1993; Kim 1998). Appeal to strong or spooky emergence, on the other hand, justifiably arouses suspicion. Indeed, it is unclear why properties that emerge in a spooky fashion should be thought of as higher-level at all. Perhaps the very idea of spooky emergence is incoherent.
6.3.4 Mechanistic levels are not causally related to one another
As with levels of realization, many common assumptions about the nature of causation would appear to make causal relations between mechanistic parts and the properties or behaviors of wholes suspect. Items at different levels of aggregation and at different levels of mechanisms are intimate with one another in much the same way that items at different levels of realization are intimate. Lewis is explicit. If C causes E:
C and E must be distinct events [if they are to be causally related]—and distinct not only in the sense of nonidentity but also in the sense of nonoverlap and non-implication. It won’t do to say that my speaking this sentence causes my speaking this sentence; or that my speaking the whole of it causes my speaking the first half of it; or that my speaking causes my speaking it loudly, or vice versa. (Lewis 2000, p. 78)
The relevant kind of intimacy for levels of mechanisms is overlap between token events or processes. The relationship between LTP and the opening of NMDA receptors during LTP induction is directly analogous to the relationship between speaking the whole of a sentence and speaking its first half. The induction of LTP is partly constituted by the opening of the NMDA receptor. The would-be cause in this top-down causal claim already contains the would-be effect within it. There is nothing additional to be produced in the effect; the occurrence of the effect includes the occurrence of the cause.
What about the bottom-up case? We might say that the spark plugs cause the engine to run, all the while acknowledging that the sparking of spark plugs is part of the operation of the engine. The naturalness of this locution is at least partly due to an ambiguity in the way we commonly describe the behavior of a mechanism as a whole. Sometimes we describe it as an activity or process that starts with the mechanism’s setup conditions and ends with its termination conditions (Machamer et al. 2000). Thus we might describe Long-Term Potentiation as a process[26] or activity beginning with a rapid and repeated stimulus (called a tetanus) to the presynaptic neuron and ending with enhanced transmission across the synapse. Other times we describe the behavior of the mechanism as a whole, the phenomenon, as the product of that process (or one of its termination conditions). Thus we might say that the mechanism of Long-Term Potentiation produces a potentiated synapse. This way of speaking leads us to think in terms of the antecedent causes of potentiation: the tetanus is a distal cause, and the subsequent changes in the NMDA receptor are more proximal. If we think about the behavior of the mechanism in the second way, as a product, it is natural to think of the opening of the NMDA receptor as a cause of the synapses being potentiated (and indeed it is). But if we think about the behavior of the mechanism in the first way, as an input-output relation starting with the tetanus and ending with a potentiated synapse, then it is wrong to think of the tetanus or the opening of the NMDA receptor as a cause of that. The NMDA receptor is a part of that causal process. These are two equally acceptable ways of describing the relationship between a mechanism and a phenomenon; they are easily translated into one another. However, if one is careless, these ways of speaking and writing invite equivocation of precisely the sort that we are struggling here to avoid.
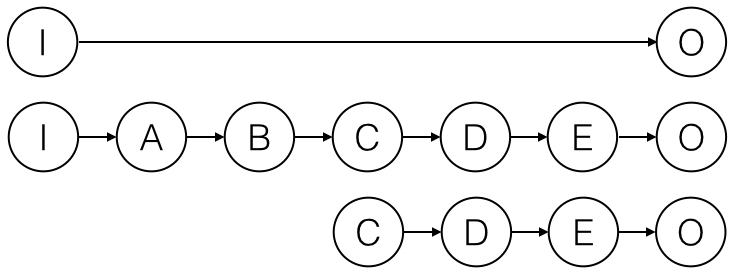 Figure 5: Why bottom-up causation is conceptually problematic.
Figure 5: Why bottom-up causation is conceptually problematic.
Suppose we represent the input-output relationship constituting LTP as in the top of Figure 5, where I is a tetanus and O is a stable, potentiated synapse. Beneath this abstract I-O relation is a more detailed description of the intermediate stages in this mechanism: the tetanus excites the postsynaptic cell (A) which depolarizes it (B), causes NMDA receptors to open (C), and so on. (Nothing turns on the fact that I’ve idealized this mechanism as a single, step-wise causal chain.) Now, suppose we intervene to open the NMDA receptors directly and thereby potentiate the synapse (as shown in the third line). We might say that this intervention induced LTP; but when we say this, we mean that it produces the end product of the mechanism (it potentiates the synapse). We cannot coherently assert that it causes the process as depicted in the first two lines. This is for the simple reason that the process in the first two lines includes stages I, A, and B, and these are absent in the causal sequence represented in the third line. At most we can say the intervention caused the last half of the process. The NMDA receptor is not a cause of the process of LTP; it is a component of that process.[27]
6.4 Levels of mechanisms in relation to other kinds of levels
This application of the levels metaphor, according to which levels of organization are understood in terms of levels of aggregation and levels of mechanisms, thus offers a no-nonsense, ontological picture that comports well with the kinds of explanatory structure one finds in neuroscience and throughout the special sciences generally.
This view eschews the idea that levels are monolithic strata in the structure of the universe, with proprietary causal laws and forces (contra the view in Wimsatt, Oppenheim, and Putnam). Likewise, it allows that items at higher levels have causal powers that things at lower levels do not, in contrast to levels of realization. Single sciences and theories might investigate phenomena at many levels of organization, and an item located at one level in such a compositional hierarchy might be studied by many sciences and described in many theories. Things at different levels of organization (aggregation and mechanism) do not causally interact with one another, though we might find more complicated ways of describing how these items depend upon one another (see Craver & Bechtel 2007). As a result, if we think about the world in terms of levels of organization, we should not be tempted into thinking that things at higher levels control or dominate things at lower levels. Levels of organization are, in a sense, levels of explanation, given that explanations for different topping off phenomena will often decompose the system into altogether different parts within parts. It might be difficult to discern such levels in scientific practice, and the organization of components might be very complex, but nothing emerges from levels of mechanisms except in the banal sense that parts organized together do things that the parts alone cannot. Levels of organization, in other words, seem to capture many of the intuitions that accompany the idea that the world is organized into levels but without many of the objectionable elements of other applications of the levels metaphor. The fact that the levels metaphor is often used carelessly and deployed in ways that violate common sense and metaphysical ideas about the structure of the world should not lead one to abandon the metaphor entirely. As we’ve seen, it can be given a relatively precise and metaphysically unobjectionable formulation that, in addition, fits the multilevel structures that the most advanced special sciences seem to be discovering.