3 The vestibular system
In this section, we describe the basic mechanisms of the peripheral and central vestibular system for coding self-motion and self-orientation, as we believe that these aspects are crucial bases for a sense of the bodily self. It is, however, beyond the scope of this paper to provide a comprehensive description of the vestibular system anatomy and physiology, and the reader is referred to recent review articles (e.g., Angelaki & Cullen 2008; Lopez & Blanke 2011).
3.1 Peripheral mechanisms
The peripheral vestibular organs in the inner ear contain sensors detecting three-dimensional linear motions (two otolith organs) and angular motions (three semicircular canals). The characteristic of these sensors is that they are inertial sensors, a type of accelerometers and gyroscopes found in inertial navigation systems. When an individual turns actively his or her head, or when the head is moved passively (e.g., in a train moving forward), the head acceleration is transmitted to the vestibular organs. Head movements create inertial forces—due to the inertia of the otoconia, the small crystals of calcium carbonate above the otolith organs, and to the inertia of the endolymphatic fluid in the semicircular canals—inducing an activation or inactivation of the vestibular sensory hair cells.
It is important to note here that the neural responses of the vestibular sensory hair cells depend on the direction of head movements with respect to head-centred inertial sensors and not with respect to any external reference. For this reason, the vestibular system enables the coding of absolute head motion in a head-centred reference frame (Berthoz 2000). This way of coding body motion differs from the motion coding done by other sensory systems. The coding by the visual, somatosensory, and auditory system is ambiguous because these sensory systems detect a body motion relative to an external reference, or the motion of an external object with respect to the body. For example, the movement of an image on the retina can be interpreted either as a motion of the body with respect to the visual surrounding, or as a motion of the visual scene in front of a static observer (e.g., Dichgans & Brandt 1978), leading to an ambiguous sense of ownership for the movement. Similarly, if a subject detects changes of pressures applied to his skin (e.g., under his foot soles), this can be related either to a body movement, with respect to the surface on which he is standing, or to the movement of this surface on his skin (Kavounoudias et al. 1998; Lackner & DiZio 2005). Similar observations have been made in the auditory system and illusory sensations of body motion have been evoked by rotating sounds (Väljamäe 2009). By contrast, a vestibular signal is a non-ambiguous neural signal that the head moved or has been moved; thus there is no ambiguity regarding whether the own body moved or the environment moved. It should, however, be noted that the vestibular information on its own does not distinguish between passive or active movements of the subject’s whole body (i.e., the self-motion associated with the feeling of agency; see also section 4.4).[8]
The otolith organs are not only activated by head translations, such as those produced by a train moving forward or by an elevator moving upward, but also by Earth’s gravitational pull. Otolith receptors are sensitive to gravito-inertial forces (Angelaki et al. 2004; Fernández & Goldberg 1976) and thus provide the brain with signals about head orientation with respect to gravity. Such information is crucial to maintain one’s body in a vertical orientation and to orient oneself in the physical world (Barra et al. 2010).
3.2 Central mechanisms
The vestibulo-thalamo-cortical pathways that transmit vestibular information from the peripheral vestibular organs to the cortex involve several structures relaying and processing vestibular sensory signals. We describe below vestibular sensory processing in the vestibular nuclei complex, thalamus, and cerebral cortex.
3.2.1 The vestibular nuclei complex and thalamus
The eighth cranial nerve transmits vestibular signals from the vestibular end organs to the vestibular nuclei complex and cerebellum (Barmack 2003). The vestibular nuclei complex is located in the brainstem and is the main relay station for vestibular signals. From the vestibular nuclei, descending projections to the spinal cord are responsible for vestibulo-spinal reflexes and postural control. Ascending projections to the oculomotor nuclei support eye movement control, while ascending projections to the thalamus and subsequently to the neocortex support the vestibular contribution to higher brain functions. Vestibular nuclei are also strongly interconnected with several nuclei in the brainstem and limbic structures, enabling the control of autonomic functions and emotion (see section 4.1.3) (Balaban 2004; Taube 2007).
The role of the vestibular nuclei is not limited to a relay station for vestibular signals. Complex sensory processing takes place in vestibular nuclei neurons, involving, for example, the distinction between active, self-generated head movements and passive, externally imposed head movements (Cullen et al. 2003; Roy & Cullen 2004). As we will argue in section 4.4, this processing is likely to play a crucial role in the sense of agency, especially concerning full-body agency during locomotion. Another characteristic of the vestibular nuclei complex is the large extent of multisensory convergence that occurs within it (Roy & Cullen 2004; Tomlinson & Robinson 1984; Waespe & Henn 1978), which leads to the perceptual “disappearance” of vestibular signals as they are merged with eye movement, visual, tactile, and proprioceptive signals. Because there is “no overt, readily recognizable, localizable, conscious sensation” from the vestibular organs during active head movements, excluding artificial passive movements and pathological rotatory vertigo, the vestibular sense has been termed a “silent sense” (Day & Fitzpatrick 2005).
Ascending projections from the vestibular nuclei complex reach the thalamus. These projections are bilateral and very distributed as there is no thalamic nucleus specifically dedicated to vestibular processing, as compared to visual, auditory, or tactile processing.[9] Anatomical and electrophysiological studies in rodents and primates identified vestibular neurons in many thalamic nuclei (review in Lopez & Blanke 2011). Important vestibular projections have been noted in the ventroposterior complex of the thalamus, a group of nuclei typically involved in somatosensory processing (Marlinski & McCrea 2008a; Meng et al. 2007). Other vestibular projections have been identified in the ventroanterior and ventrolateral nuclear complex, intralaminar nuclei, as well as in the lateral and medial geniculate nuclei (Kotchabhakdi et al. 1980; Lai et al. 2000; Meng et al. 2001). Electrophysiological studies revealed that similarly to vestibular nuclei neurons, thalamic vestibular neurons can distinguish active, self-generated head movements from passive head movements, showing a convergence of vestibular and motor signals in the thalamus (Marlinski & McCrea 2008b).
3.2.2 Vestibular projections to the cortex
Vestibular processing occurs in several cortical areas as demonstrated as early as the 1940s in the cat neocortex and later in the primate neocortex (reviews in Berthoz 1996; Fukushima 1997; Grüsser et al. 1994; Guldin & Grüsser 1998; Lopez & Blanke 2011). Figure 3 summarizes the main vestibular areas found in the monkey and human cerebral cortex. More than ten vestibular areas have been identified to date.
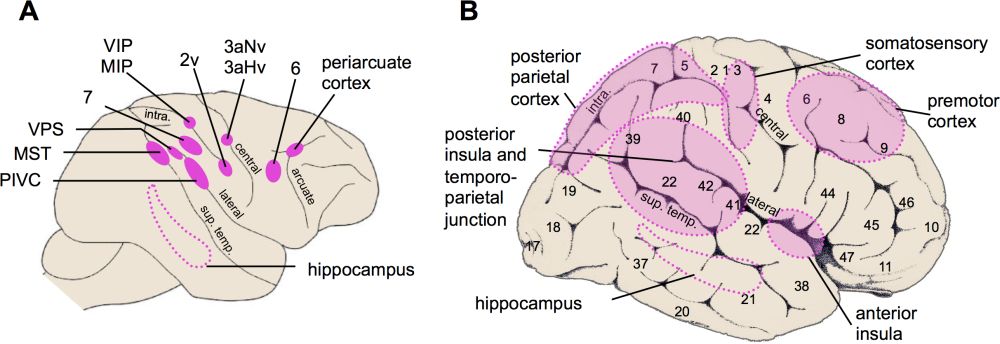 Figure 3: Schematic representation of the main cortical vestibular areas. (A) Main vestibular areas in monkeys are somatosensory areas 2v and 3av (3aHv (3a-hand-vestibular region), 3aNv (3a-neck-vestibular region)) in the postcentral gyrus, frontal area 6v and the periarcuate cortex, parietal area 7, MIP (medial intraparietal area) and VIP (ventral intraparietal area), extrastriate area MST (medial superior temporal area), PIVC (parieto-insular vestibular cortex), VPS (visual posterior sylvian area), and the hippocampus. Major sulci are represented: arcuate sulcus (arcuate), central sulcus (central), lateral sulcus (lateral), intraparietal sulcus (intra.), and superior temporal sulcus (sup. temp.). Adapted from Lopez and Blanke after Sugiuchi et al. (2005). (B) Main vestibular areas in the human brain identified by noninvasive functional neuroimaging techniques. Numbers on the cortex refer to the cytoarchitectonic areas defined by Brodmann. Adapted from Lopez & Blanke (2011) after Sugiuchi et al. (2005).
Figure 3: Schematic representation of the main cortical vestibular areas. (A) Main vestibular areas in monkeys are somatosensory areas 2v and 3av (3aHv (3a-hand-vestibular region), 3aNv (3a-neck-vestibular region)) in the postcentral gyrus, frontal area 6v and the periarcuate cortex, parietal area 7, MIP (medial intraparietal area) and VIP (ventral intraparietal area), extrastriate area MST (medial superior temporal area), PIVC (parieto-insular vestibular cortex), VPS (visual posterior sylvian area), and the hippocampus. Major sulci are represented: arcuate sulcus (arcuate), central sulcus (central), lateral sulcus (lateral), intraparietal sulcus (intra.), and superior temporal sulcus (sup. temp.). Adapted from Lopez and Blanke after Sugiuchi et al. (2005). (B) Main vestibular areas in the human brain identified by noninvasive functional neuroimaging techniques. Numbers on the cortex refer to the cytoarchitectonic areas defined by Brodmann. Adapted from Lopez & Blanke (2011) after Sugiuchi et al. (2005).
Electrophysiological and anatomical studies in animals have revealed important vestibular projections to a region covering the posterior parts of the insula and lateral sulcus, an area referred to as the parieto-insular vestibular cortex (PIVC) (Grüsser et al. 1990a; Guldin et al. 1992; Liu et al. 2011). Other vestibular regions include the primary somatosensory cortex (the hand and neck somatosensory representations of postcentral areas 2 and 3 [Ödkvist et al. 1974; Schwarz et al. 1973; Schwarz & Fredrickson 1971]); ventral and medial areas of the intraparietal sulcus (Bremmer et al. 2001; Chen et al. 2011; Schlack et al. 2005); visual motion sensitive area MST (Bremmer et al. 1999; Gu et al. 2007); frontal cortex (motor and premotor cortex and the frontal eye fields [Ebata et al. 2004; Fukushima et al. 2006]); cingulate cortex (Guldin et al. 1992) and hippocampus (O’Mara et al. 1994). These findings indicate that vestibular processing in the animal cortex relies on a highly distributed cortical network.
A similar conclusion has been drawn from neuroimaging studies conducted in humans. These studies have used fMRI and PET during caloric and galvanic vestibular stimulation[10] and revealed that the human vestibular cortex closely matches the vestibular regions found in animals. Vestibular responses were found in the insular cortex and parietal operculum as well as in several regions of the temporo-parietal junction (superior temporal gyrus, angular and supramarginal gyri). Other vestibular activations are located in the primary and secondary somatosensory cortex, precuneus, cingulate cortex, frontal cortex, and hippocampus (Bense et al. 2001; Bottini et al. 1994; Bottini et al. 1995; Dieterich et al. 2003; Eickhoff et al. 2006; Indovina et al. 2005; Lobel et al. 1998; Suzuki et al. 2001).
It is of note that the non-human animal and human vestibular cortex differs from other sensory cortices as there is apparently no primary vestibular cortex; that is, there is no koniocortex dedicated to vestibular processing and containing only or mainly vestibular responding neurons (Grüsser et al. 1994; Guldin et al. 1992; Guldin & Grüsser 1998), stressing again the multisensory character of the vestibular system. All areas processing vestibular signals are multimodal, integrating visual, tactile, and proprioceptive signals. The PIVC has been shown to occupy a key role in the cortical vestibular network and is the only vestibular area that is connected to all other vestibular regions described above. The PIVC also receives signals from the primary somatosensory cortex, premotor cortex, posterior parietal cortex, and the cingulate cortex (Grüsser et al. 1994; Guldin et al. 1992), and it integrates signals from personal and extrapersonal spaces. Given these characteristics, we believe that the PIVC should be importantly involved in a coherent representation of the bodily self and the body embedded in the world.
3.2.3 The PIVC as a core, multimodal, vestibular cortex
The group of Grüsser was the first to describe vestibular responses in the monkey PIVC. Vestibular neurons were located in several regions of the posterior end of the lateral sulcus “in the upper bank of the lateral sulcus around the posterior end of the insula, sometimes also within the upper posterior end of the insula [… and] more posteriorly in the retroinsular region or more anteriorly in the parietal operculum” (Grüsser et al. 1990a, pp. 543-544; Grüsser et al. 1990b; Guldin et al. 1992; Guldin & Grüsser 1998). Figure 4A illustrates the location of PIVC in the macaque brain. Recent investigations of PIVC in rhesus monkeys revealed that vestibular neurons were mostly located in the retroinsular cortex and at the junction between the secondary somatosensory cortex, retroinsular cortex, and granular insular cortex (area Ig) (Chen et al. 2010; Liu et al. 2011).
In humans, functional neuroimaging studies used caloric and galvanic vestibular stimulation and showed activations in and around the posterior insula and temporo-parietal junction (Bense et al. 2001; Bottini et al. 1994; Dieterich et al. 2003; Eickhoff et al. 2006; Lobel et al. 1998; Suzuki et al. 2001). Because these activations also extend to the superior temporal gyrus, posterior and anterior insula, and inferior parietal lobule, the exact location of human PIVC is still debated (review in Lopez & Blanke 2011). Recent meta-analyses of vestibular activations suggest that the core vestibular cortex is in the parietal operculum, retroinsular cortex, and/or posterior insula (Lopez et al. 2012; zu Eulenburg et al. 2012) (figure 4B). Of note, several neuroimaging studies have also implicated the anterior insula in vestibular processing (Bense et al. 2001; Bottini et al. 2001; Fasold et al. 2002). The insula is crucial for interoceptive awareness (Craig 2009) and could provide the neural substrate for vestibulo-interoceptive interactions that impact several aspects of the bodily self (see section 4.1.3).
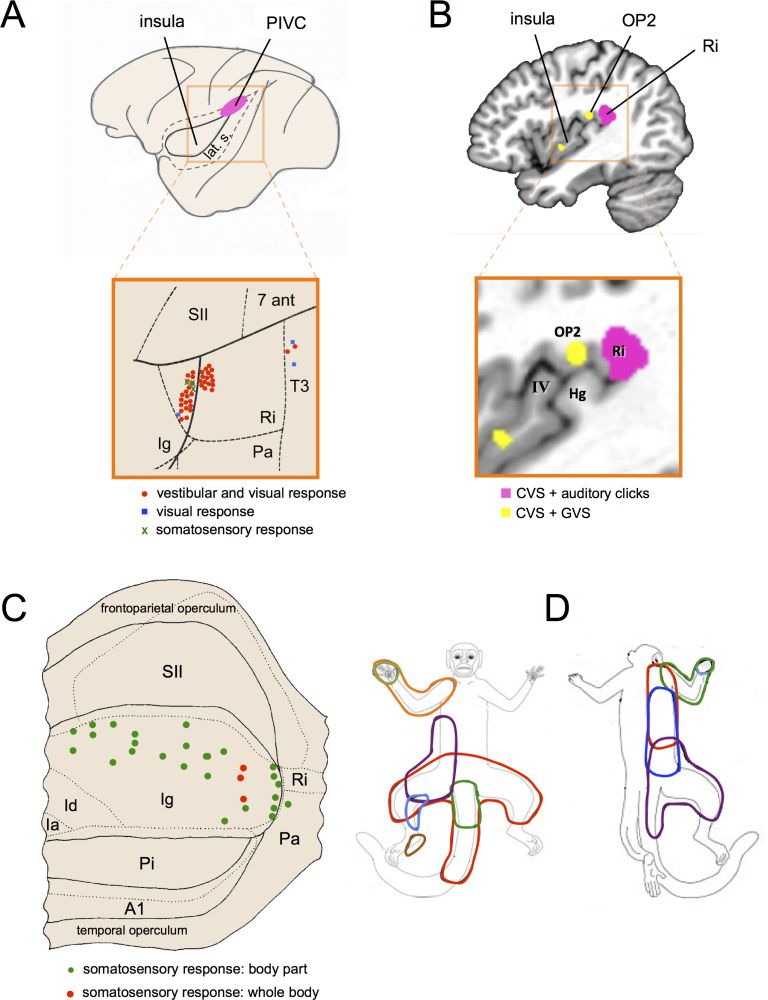 Figure 4: Anatomical location and functional properties of the parieto-insular vestibular cortex (PIVC). (A) Schematic representation of the macaque brain showing the location of the PIVC. For the purpose of illustration, the lateral sulcus (lat. s.) is shown unfolded. The macaque PIVC is located in the parietal operculum at the posterior end of the insula and retroinsular cortex. Modified from Grüsseret al. (1994). The insert illustrates the location of vestibular neurons in different regions of the lateral sulcus in a squirrel monkey (Saimiri sciureus). The lateral sulcus is shown unfolded to visualize the retroinsular cortex (Ri), secondary somatosensory cortex (SII), granular insular cortex (Ig), and auditory cortex (PA). Vestibular neurons (red dots) were mostly located in Ri and Ig. Adapted from Guldin et al. (1992). (B) Vestibular activations found in the human PIVC using meta-analysis of functional neuroimaging data. The Ri showed a convergence of activations evoked by caloric vestibular stimulation (CVS) of the semicircular canals and auditory activation of the otolith organs (pink). The parietal operculum (OP2) and posterior insula showed a convergence of activations evoked by CVS and galvanic vestibular stimulation (GVS) of all primary vestibular afferents (yellow). Hg (Heschl’s gyrus). Adapted from Lopez et al. (2012). (C) View of the unfolded lateral sulcus of the rhesus monkey (Macaca mulatta) showing somatosensory neurons (green dots) in the granular insula, of which some have large somatosensory receptive fields covering the whole body (red dots). Ia (agranular insular field); Id (dysgranular insular field); A1 (first auditory field); Pa (postauditory field); Pi (parainsular field). Modified from Schneider et al. (1993). (D) Representation of the size of the receptive fields of neurons recorded in somatosensory representations of the body found in the dorsal part of the insula (ventral somatosensory area) of the titi monkey (Callicebus moloch). Modified from Coq et al. (2004).
Figure 4: Anatomical location and functional properties of the parieto-insular vestibular cortex (PIVC). (A) Schematic representation of the macaque brain showing the location of the PIVC. For the purpose of illustration, the lateral sulcus (lat. s.) is shown unfolded. The macaque PIVC is located in the parietal operculum at the posterior end of the insula and retroinsular cortex. Modified from Grüsseret al. (1994). The insert illustrates the location of vestibular neurons in different regions of the lateral sulcus in a squirrel monkey (Saimiri sciureus). The lateral sulcus is shown unfolded to visualize the retroinsular cortex (Ri), secondary somatosensory cortex (SII), granular insular cortex (Ig), and auditory cortex (PA). Vestibular neurons (red dots) were mostly located in Ri and Ig. Adapted from Guldin et al. (1992). (B) Vestibular activations found in the human PIVC using meta-analysis of functional neuroimaging data. The Ri showed a convergence of activations evoked by caloric vestibular stimulation (CVS) of the semicircular canals and auditory activation of the otolith organs (pink). The parietal operculum (OP2) and posterior insula showed a convergence of activations evoked by CVS and galvanic vestibular stimulation (GVS) of all primary vestibular afferents (yellow). Hg (Heschl’s gyrus). Adapted from Lopez et al. (2012). (C) View of the unfolded lateral sulcus of the rhesus monkey (Macaca mulatta) showing somatosensory neurons (green dots) in the granular insula, of which some have large somatosensory receptive fields covering the whole body (red dots). Ia (agranular insular field); Id (dysgranular insular field); A1 (first auditory field); Pa (postauditory field); Pi (parainsular field). Modified from Schneider et al. (1993). (D) Representation of the size of the receptive fields of neurons recorded in somatosensory representations of the body found in the dorsal part of the insula (ventral somatosensory area) of the titi monkey (Callicebus moloch). Modified from Coq et al. (2004).